近日,上海师范大学杨洪全教授团队在The Plant Cell在线发表了题为Arabidopsis Cryptochrome 1 Controls Photomorphogenesis through Regulation of H2A.Z Deposition的研究论文,该论文揭示了拟南芥蓝光受体隐花素CRY1通过调控H2A.Z占位来调控光形态建成的分子机制。
太阳光不仅是植物进行光合作用的能量来源,同时也是植物生长发育调控的重要信号来源。植物可以通过多种光受体来感受不同波长、强度和方向的光,从而来适应不同光环境下进行生长和发育。蓝光受体隐花素(cryptochrome)是最早被发现的蓝光受体,在拟南芥中有2个CRY:CRY1和CRY2。CRY1主要参与调控光形态建成,而CRY2则主要参与调控光周期开花时间。CRY1蛋白通过其的C端延伸结构域可以与E3泛素连接酶COP1以及COP1的增强子SPAs蛋白发生相互作用,抑制COP1-SPAs复合物的活性,促进COP1的底物------光形态建成的正调控因子HY5的蛋白积累,从而促进光形态建成。HY5作为bZIP家族转录因子可以促进光形态建成相关基因表达(例如参与叶绿素合成和光合作用的基因),抑制暗形态建成相关基因的表达(例如促进下胚轴伸长过程的基因)。
H2A.Z是进化上非常保守的组蛋白H2A的变体蛋白,从最简单的真核生物酿酒酵母到人类中都广泛分布,参与了转录调控和DNA修复等重要过程。H2A.Z在染色质上的替换主要是由ATP依赖的SWR1复合物来催化完成,在拟南芥中研究比较清楚的复合物亚基是SWC6和ARP6,两者存在相互作用,并且都位于复合物的C模块。杨洪全教授实验室以CRY1的N端功能区(CNT1)作为诱饵蛋白,通过酵母双杂交系统筛选相互作用蛋白,获得了一个新的CRY1的互作蛋白SWC6。通过一系列蛋白—蛋白相互作用的实验分析,发现CRY1分别与SWC6和ARP6发生蓝光依赖性的互作。通过突变体下胚轴表型分析,发现SWC6、ARP6和H2A.Z在蓝光、红光和远红光下都抑制下胚轴的伸长,在蓝光下还可以促进叶绿素的合成。通过转录组测序的方法,分析了CRYs和ARP6在蓝光下共同调控的基因,其中有73%的基因被两者同向调控;通过分析ARP6和HY5/HYH共同调控基因,发现其中有88%的基因被两者同向调控。RT-qPCR分析显示,很多促进下胚轴伸长基因的表达同时被CRYs、ARP6和HY5/HYH所抑制,而一些促进叶绿素合成和参与光合作用相关的基因表达则同时被CRYs、ARP6和HY5/HYH所促进。通过ChIP-qPCR分析,发现HY5的靶基因EXP2、IAA19、XTH33、HEMA1和RBCS1A上的H2A.Z的占位依赖于SWC6和ARP6。进一步的ChIP-qPCR分析表明,CRYs介导蓝光促进EXP2、IAA19和XTH33基因上的H2A.Z占位,以及HY5/HYH促进这些基因上的H2A.Z的占位。通过蛋白—蛋白互作分析,发现HY5和HYH分别与SWC6和ARP6发生直接互作,由此可能招募SWR1复合物来促进H2A.Z的占位。进一步通过蛋白互作分析,发现蓝光激活的CRY1可以促进SWC6和ARP6之间的互作,进而可能促进SWR1复合物的活性。这些结果表明,在蓝光条件下,CRY1一方面可能通过与SWC6和ARP6互作,来增强SWR1复合物活性来促进H2A.Z的占位;另一方面还可能通过抑制COP1的活性来促进HY5蛋白的积累,通过与SWC6/ARP6的互作来增加对SWR1复合物的招募量来促进H2A.Z的占位。上述两层调控双管齐下,CRY1可以高效介导蓝光信号对H2A.Z在HY5靶基因上占位的促进,从而促进光形态建成。结合这两方面的因素,CRY1可以促进H2A.Z在HY5靶基因(促进下胚轴伸长)上的H2A.Z占位,抑制这些基因的表达,从而促进光形态建成过程。另外,这些结果暗示SWC6和ARP6是HY5潜在的伴侣蛋白,来参与调控HY5的转录活性。
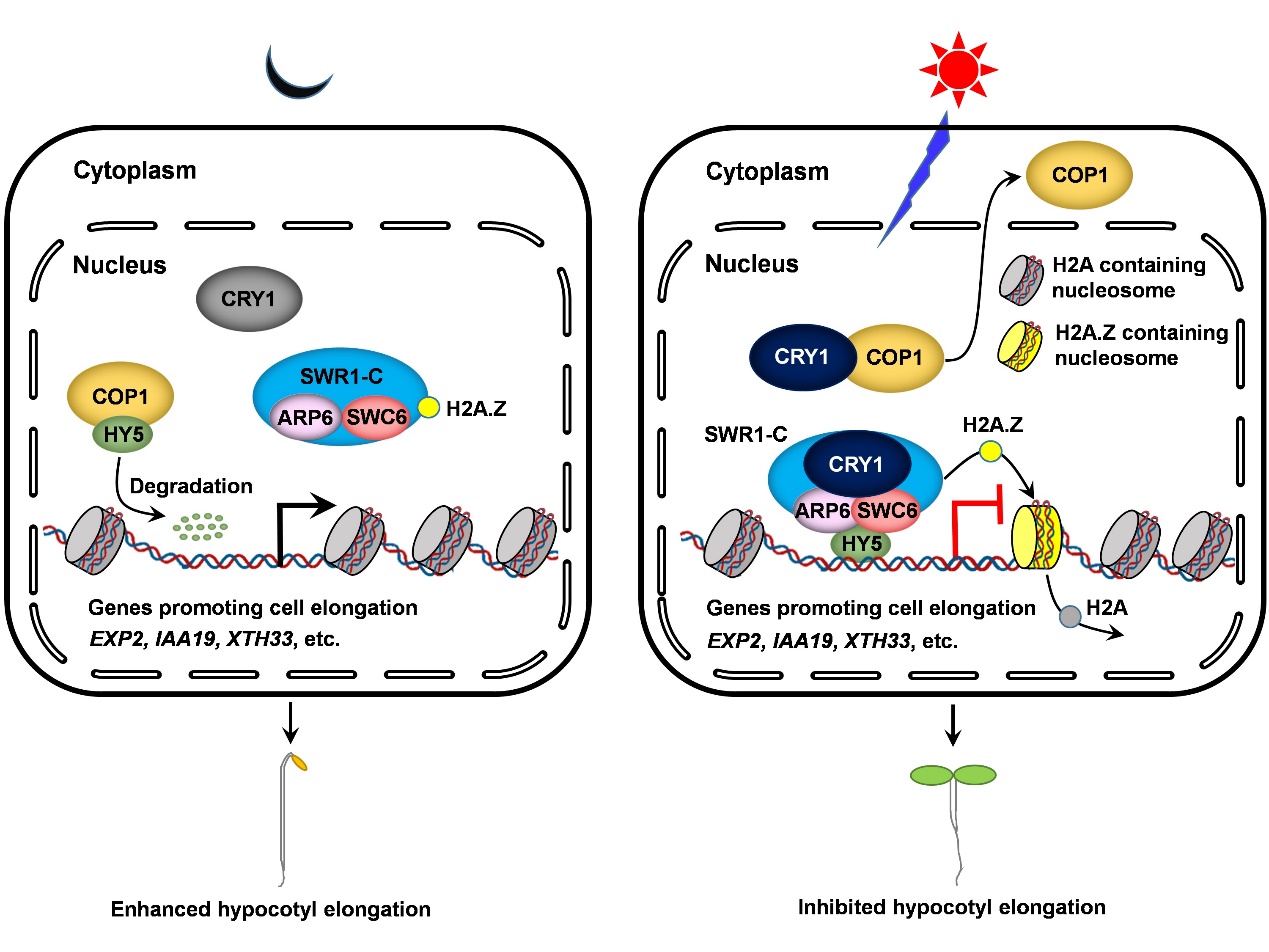
蓝光受体CRY1通过促进H2A.Z占位来促进植物光形态建成机制的模型图
该研究工作的第一作者为上海师范大学的茅志磊博士,通讯作者为杨洪全教授。该工作得到了科技部、国家自然科学基金委和上海市重点实验室项目的资助。
论文链接:https://doi.org/10.1093/plcell/koab091

